BIOLOGICAL ACTION SPECTRA
Holly L. Gorton
Department of Biology
St. Mary's College of Maryland
St. Mary's City, MD 0268
hlgorton@smcm.edu
The study of photobiology is interdisciplinary, with light being the unifying theme among such diverse fields as photosynthesis, vision, phototropism, sunburn, and low level light therapy (LLLT). Photobiologists studying different systems depend on similar photochemical and photobiological principles, and similar questions arise about vastly different light-controlled biological systems. For example, light is doing something, but what light? A central question is what wavelengths of light elicit a particular response, and the goal of action spectroscopy is to answer that question. An action spectrum is simply a plot of biological effectiveness as a function of the wavelength of incident light, and it can provide fundamental information about the system under study.
Why Measure Action Spectra?
Classically the goal of making an action spectrum was to help identify the process's photoreceptor pigment, which is often the first step in understanding the whole chain of events from absorption to signal transduction and amplification to one or more responses. The concept behind using action spectra to help identify photoreceptor pigments is simple. The First Law of Photochemistry (the Grotthus-Draper Law) states that light must be absorbed in order to have an effect, but not all light is absorbed equally. The absorption spectrum of the pigment describes how strongly different wavelengths of light are absorbed. Because more absorption leads to more action, peaks in an absorption spectrum will have corresponding peaks in an action spectrum. Thus, if a pigment can be found that has an absorption spectrum that matches a process's action spectrum, it is likely that pigment is the photoreceptor for that process. Although the concept is simple, obtaining action spectra that correspond to absorption spectra is a challenge.
Many reviews and book chapters detail the conditions that must be met to obtain a true action spectrum, i.e., one that matches the photoreceptor's absorption spectrum. Authors generally agree about these conditions; they are the same no matter what experimental system is under investigation (Setlow, 1957; Jagger, 1967; Kleczkowski, 1972; Shropshire, 1972; French, 1977; Coohill, 1984; Schäfer & Fukshansky, 1984; Sliney, 2006; Björn, 2008). This module will describe the conditions for rigorous action spectroscopy, and will also discuss examples of action spectra obtained when the classical conditions are not all fulfilled. If action and absorbance spectra do not match, one can learn a great deal about the system by finding out why. In addition, we will discuss other uses for action spectroscopy besides identifying photoreceptor pigments, uses for which experimental requirements may be simpler.
Representative Action Spectra: Learning from the Classics
Characterizing the light that effects a biological change is an obvious thing to do, so it is not surprising that scientists have used some form of action spectroscopy since the early days of experimental science, but it has taken scientists a long time to appreciate the subtleties of the measurement. We will discuss the development of action spectroscopy through representative historical examples, which any student of photobiology should know. In each case, the goal of the experiment was to help identify photoreceptor pigments. Each of these examples will inform our analysis of how action spectra should best be performed.
When first considering the effectiveness of different wavelengths of light, one might be tempted simply to shine different colors of light on the subject, and evaluate the resulting response. That is a common strategy seen in middle-school science-fair projects, and it is what the earliest steps toward action spectroscopy involved. For example, in the early 1800s, Daubeny investigated what colors of light might influence a range of plant responses, including photosynthesis, diurnal leaf movements, greening of leaves, and transpiration (Daubeny, 1836). He used colored glass filters, as well as bottles of colored solutions such as copper sulfate or, more interestingly, port wine. His methods were rudimentary, but he did provide simple transmittance spectra of his filters, and he quantified how much light came through each by measuring how much each color treatment would raise the temperature of a thermometer with a blackened bulb. Although Daubeny tried to measure how much light he was giving, he did not control it in any way. Clearly, with Daubeny's approach, the amount of light given at each wavelength must be the same if the action spectrum is to be meaningful.
The earliest action spectrum that survives in today's biology textbooks is Engelmann's photosynthesis action spectrum (Engelmann, 1882; Drews, 2005). The work endures in today's texts largely because of Engelmann's exceptionally ingenious experimental design. He modified a microscope with a prism so that light from a gaslight source was projected as a microspectrum across the microscope slide, covering about three long cells of his subject, the filamentous green alga, Cladophora. Engelmann's goal was to determine which colors of light produced the most photosynthesis, but it's not easy to see that process under a microscope. Engelmann knew that photosynthesis produced oxygen, and his trick was to use motile bacteria that were attracted to oxygen in order to visualize where photosynthetic oxygen evolution was occurring in the Cladophora filaments. His visual assay consisted of observing what colors of light caused the greatest accumulation of the aerotactic bacteria around the periphery of the alga (Figure 1). He noted that the bacteria accumulated along regions of the algal filament that were illuminated by red or blue light, and he concluded that those spectral regions were the most effective in stimulating photosynthesis in Cladophora. Most importantly, he concluded that the green pigment chlorophyll, which absorbs red and blue light most strongly, was involved in the photosynthetic process. He extended his work with similarly obtained photosynthetic action spectra for brown, red, and blue-green algae, and his results gave evidence for participation of the different photosynthetic accessory pigments in these taxonomically distinct algal groups (Engelmann, 1884). We know these pigments today as fucoxanthin, phycoerythrin, and phycocyanin.
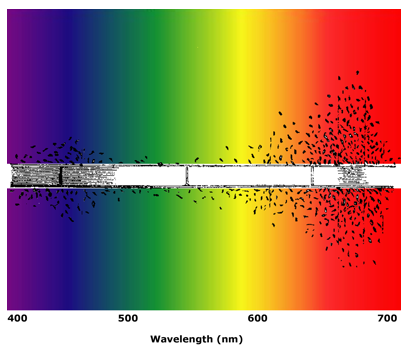
Figure 1. Engelmann's photosynthetic action spectrum. Aerotactic bacteria move toward areas of high oxygen concentration indicative of photosynthesis, stimulated by red and blue light. The black and white drawing is from (Engelmann, 1882), and it has been superimposed on a spectrum to show photosynthetically active wavelengths.
For an action spectrum like Engelmann's to make sense, one must assume that there were equal amounts of light in each part of the spectrum, but in Engelmann's case this assumption would have been incorrect. The gaslight he used would have emitted more red than blue photons, which may explain why he saw much more bacterial accumulation in the red region of the spectrum. Nonetheless, the broad pattern he observed for photosynthetic activity in the red and blue correctly led to the identification of chlorophyll as a major photosynthetic pigment. Engelmann's action spectra are crude by today's standards, but they were the first with good spectral resolution, and his conclusions remain valid.
Even if Daubeny and Engelmann were able to give equal amounts of light at all wavelengths, this method for measuring action spectra is problematic. In order to understand why this is so, consider how a photobiological response varies with the amount of light. In many, but certainly not all, photobiological systems, the magnitude of the response is proportional to the log of the amount of light. This concept, called the Weber-Fechner law, was articulated in the 19th century for human perception of various stimuli, including light. This relationship stems from signal transduction and amplification, which allow sensory systems in animals and plants to be sensitive over many orders of magnitude of light stimulus. Consider a hypothetical photoreceptor pigment with an absorption maximum at 550 nm. Figure 2 shows how a response mediated by this pigment varies logarithmically with fluence at each wavelength. Fluence is a measurement of light coming from all directions using a spherical detector. Typically it is calculated from the energy fluence rate (Wm-2) or photon fluence rate (mol m-2s-1) by the duration of the irradiation, yielding energy fluence (Jm-2) or photon fluence (µmol m-2). For more information, see the PDF file on Radiometric Quantities and Units Used in Photobiology in the section on Photophysics.
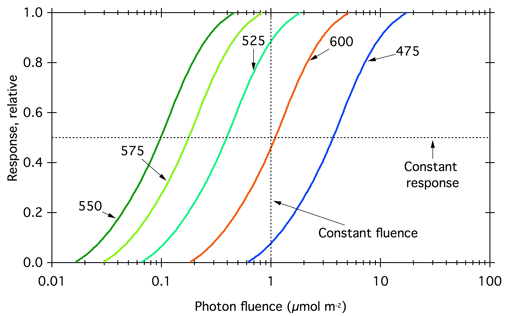
Figure 2. Fluence response curves for a hypothetical photoresponse. The curves in are typically sigmoidal, showing a threshold, a linear portion, and saturation. Dashed lines represent constant fluence and constant response used to generate the action spectra in Figure 3.
The strategy of measuring the response to equal fluences of light at different wavelengths can give different results depending on the fluence chosen. Consider the fluence-response curves in Figure 2, and suppose that the investigator measured the response to 1 µmol m-2 at each wavelength. That fluence is above saturation for 550 and 575 nm, and close to saturation for 525 nm. Plotting the response to this fluence as a function of wavelength would lead to artificial flattening of the peaks of the action spectrum (Figure 3, red curve).
A better procedure is to plot the fluence necessary to generate a constant response; wavelengths that are less strongly absorbed require more light to give the same response as wavelengths that are more strongly absorbed. Today it is common to plot action spectra as the reciprocal of the fluence required to give a particular level of response, generally 50% of the saturated response (Figure 3, blue curve). For each wavelength, the reciprocal of the fluence required to give a constant response should be proportional to the absorption coefficient of the pigment at that wavelength (Shropshire, 1972). In addition, plotting the reciprocal of the required fluence gives a curve that matches our intuitive expectation, with peaks for the most active wavelengths.
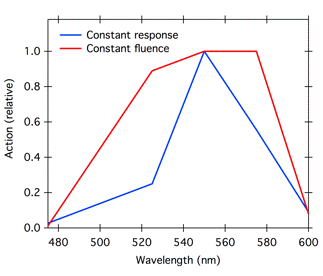
Figure 3. Action spectra for a hypothetical photoresponse constructed from the data in Figure 2. Plotting the response to equal fluence (1.0 µmol m-2) at different wavelengths generated an artificially flattened peak (red curve), because the fluence chosen was above saturation for some of the wavelengths. The blue curve represents the reciprocal of the fluence necessary to generate 50% response at each wavelength in Figure 2, and is a more accurate representation of relative action.
In the early 1900s, there was intense interest on the effects of UV irradiation on organisms, and by that time the benefits of using fluence-response curves to generate action spectra were understood. When Gates studied the bactericidal effect of UV radiation on Staphylococcus aureus, he generated fluence-response curves (Figure 4) to determine how much light was necessary to induce 50% killing at each wavelength, and plotted the reciprocal of those values as his action spectrum (Figure 5). Gates noted that his action spectrum followed the absorption spectrum of nucleic acid derivatives (Figure 5) (Gates, 1928; 1930). His action spectra provided an early indication of the possibility that DNA was the genetic material.
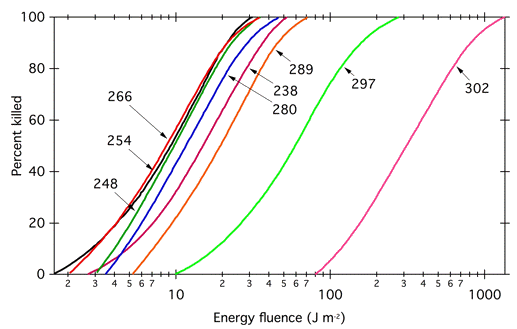
Figure 4. Fluence-response curves used to construct the action spectrum for bactericidal effects of UV radiation shown in Figure 5. Modified from (Gates, 1930). Each curve shows effectiveness for monochromatic radiation of the wavelength (nm) indicated.
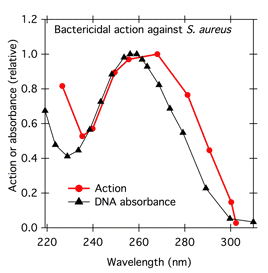
Figure 5. Action spectrum for bactericidal action of UV against Staphylococcus aureus (modified from Gates, 1930), plotted with the absorbance spectrum for DNA (modified from Tsuboi, 1950).
By the 1940s the study of the relative efficacy of different wavelengths of visible and UV radiation was common, and it had become known as action spectroscopy (Kleczkowski, 1972). Plant biologists were working to understand the red/far red photoreversible control of a number of plant responses, including seed germination, photoperiodic flowering, and stem elongation. Action spectra for red-light induction and far-red light reversal of these and other light-mediated responses were similar, suggesting that they were all mediated by the same pigment system, one we know today as the phytochrome system. By the mid 1900s, techniques for obtaining reliable action spectra that reflect the absorption spectra of photoreceptor pigments were well developed, and there are many examples of action spectra for phytochrome-mediated responses. The action spectra for red-light induction and far-red-light reversal of stem straightening (hypocotyl hook opening) in bean (Phaseolus) seedlings are classic (Figure 6).
Note that Gates's UV fluence-response curves, and hence his action spectrum expressed fluence in energy units; his focus was on how much UV energy was required to inactivate the bacteria. But recall that photochemistry occurs on a photon-by-photon basis. Thus, to aid in identifying a photoreceptor molecule, what is important is how many photons are necessary, not how much energy. The phytochrome action spectra developed by Withrow et al. (Figure 6) reflect this understanding. These sorts of spectra provided a tool with which to measure phytochrome, a prerequisite to isolation of the pigment. Today we know that the phytochrome responsible for stem straightening exists in two forms, which are interconvertible by light. Absorption of photons by the inactive, red-absorbing form of phytochrome, Pr, converts it to the active far red absorbing form, Pfr, which then triggers a physiological response. Pfr absorbs far red light most strongly, and absorption converts it back to inactive Pr. The phytochrome system is described in more detail in the module on Basic Photomorphogenesis.
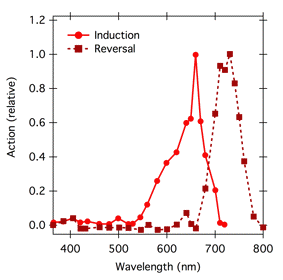
Figure 6. Action spectra for induction and reversal of hypocotyl hook opening in Phaseolus. For induction, action was calculated as the reciprocal of the photon fluence necessary to trigger a straightening of 8°. For reversal, saturating red irradiation that would have induced maximal straightening was followed by varying irradiation with different wavelengths of light. Action for reversal was calculated as the reciprocal of the photon fluence necessary to reduce the red-induced straightening by 50%. Data from (Withrow et al., 1957).
Let us consider some other examples to demonstrate the importance of using photon units rather than energy units in action spectroscopy. We will use Gates's UV action spectrum again, and also a well-constructed photosynthetic action spectrum. Because the energy per photon is inversely proportional to wavelength, the amount of distortion in an action spectrum caused by using energy units depends on the wavelength range. For example, the Gates action spectrum for the bactericidal action of UV radiation covered only 225-302 nm, so the shape of the action spectrum is not changed dramatically whether it is calculated on a photon or energy basis (Figure 7a). In contrast, action spectra covering a broader wavelength range may be more distorted. For example, an action spectrum for photosynthesis in the green alga Ulva covered 410-718 nm, and has an artificially low peak in the blue when calculated on an energy basis (Figure 7b).
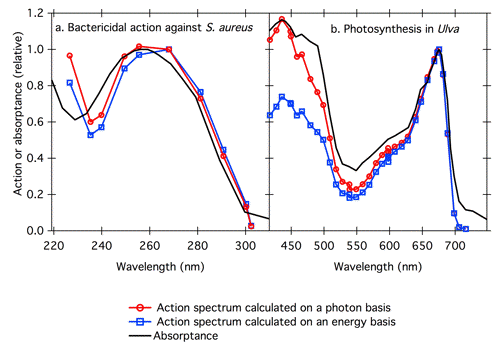
Figure 7. Action spectra for (a) bactericidal action of UV radiation against S. aureus (left), and (b) for photosynthesis in the green alga Ulva. Action spectra are shown as calculated from fluences expressed in energy units, and in photon units. For (a), the published action spectrum (Gates, 1930) was expressed in energy units (blue squares), and recalculated here on a photon basis (red circles). The solid line is an absorptance spectrum for DNA, calculated from a published absorbance spectrum (Tsuboi, 1950). For (b) the published action spectrum (Haxo & Blinks, 1950) (red circles) was calculated on a photon basis, and the data are recalculated here on an energy basis (blue squares). The solid line shows thallus (green shoot) absorptance of the alga (Haxo & Blinks, 1950). For both the bactericidal action of UV radiation, and for photosynthesis, the absorptance spectrum of the photoreceptor matches the action spectrum more closely when it is calculated on a photon basis. Note the different wavelength ranges for the two panels.
Most readers are familiar with absorbance spectra where:
Absorbance is commonly used in biochemical work, because it is proportional to concentration (see the module on Basic Photochemistry, for more information). However, note that absorptance, not absorbance, spectra are plotted in Figure 7. Absorptance is simply the fraction of incident light that is absorbed:
Because the number of photons absorbed is important, absorptance rather than the log quantity absorbance is more appropriate for comparison with action spectra. Absorptance and absorbance spectra can have very different shapes. For example, compare Figure 5, showing Gates's UV action spectrum plotted with the absorbance spectrum of DNA, with Figure 7a, showing the same action spectrum (blue squares) plotted with the corresponding absorptance spectrum (Figure 7a). The action spectrum is a somewhat closer fit to the absorptance spectrum than to the absorbance spectrum. In another example, Haxo's action spectrum for photosynthesis in Ulva fits the absorptance spectrum of the thallus better than it fits the absorbance spectrum (Figure 8). At very low concentrations of absorbing substances (less than 5% absorptance), absorptance is proportional to absorbance, and the spectra have the same shape (French, 1977).
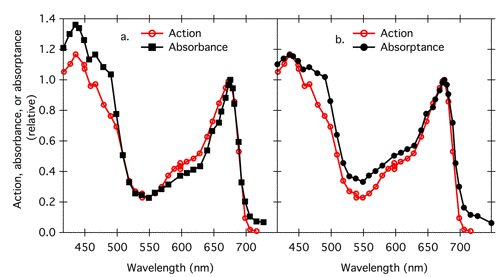
Figure 8. Action spectrum of photosynthesis in Ulva plotted with (a) thallus absorbance and (b) thallus absorptance. Action (on a photon basis) and absorptance spectra are the same as in Figure 7, modified from (Haxo & Blinks, 1950); absorbance was calculated from absorptance.
Obtaining Action Spectra: Additional Considerations.
We have seen the historical development of our current understanding of action spectra, including the accurate measurement of the light and the use of fluence-response curves, photon units and absorptance. There are a number of additional considerations relevant to the acquisition of an action spectrum that matches the absorptance spectrum of the photoreceptor pigment.
The sample should be practically transparent to all wavelengths tested (Kleczkowski, 1972). For an in vitro system, this means that the photoreceptor molecules must be diluted in a non-absorbing, stirred medium. If this condition is not met, then photoreceptor molecules might shade each other such that photons absorbed most strongly would be absorbed close to the surface while those absorbed less strongly would penetrate more deeply into the sample. Those less strongly absorbed wavelengths could still be absorbed, however, because they penetrate further, they have a longer pathlength, and the probability of absorption depends on pathlength. Thus, with increasing photoreceptor concentration, less-strongly-absorbed photons get absorbed anyway, and peaks of an action spectrum are artificially broadened. At the extreme, if the sample is so concentrated that it looks black, all photons are absorbed, and the action spectrum becomes flat over the entire wavelength range tested.
While one can manipulate pigment concentration in vitro, living systems present special challenges with respect to optical transparency. Often the photoreceptor is not close to the irradiated surface; the investigator may not even know where it is, and is unlikely to be able to control its concentration. In addition, cells and tissues may be physically thick, and thick samples present the same problems as concentrated samples described above, changing absorptance and action. Compare the absorptance spectra of isolated photosynthetic pigments, disrupted chloroplasts, whole chloroplasts, and whole leaves (Figure 9a). Light scattering, and hence pathlength, increases with increasing level of organization, and the absorptance spectrum flattens. One can see the resultant effect of structural organization on action spectra by comparing an action spectrum for photosynthesis in a green alga with that for a leaf (Figure 9b). The algal specimen is thin; green photons, which are not absorbed strongly by chlorophyll and carotenoids, pass through the thallus and out the other side without being absorbed. Absorptance, and hence action, is low for green light. In contrast, in the leaf, red and blue light are mostly absorbed in the first layer of photosynthetic cells, but green light can penetrate into the leaf interior, gaining chances to be absorbed as it travels a longer pathlength with multiple reflections at cell wall/air interfaces. Green light penetrates further into the leaf, but little escapes. The photosynthetic action spectrum for a leaf thus shows much more action in the green region of the spectrum than the corresponding action spectrum for an alga.
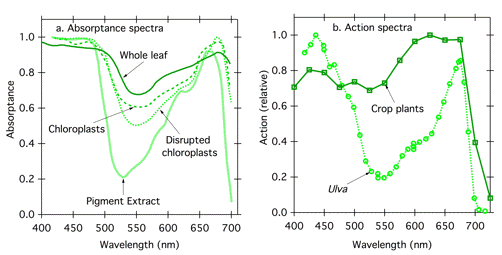
Figure 9. Effect of increasing structural complexity on absorptance and action spectra. (a) absorptance spectra of isolated pigments, disrupted chloroplasts, intact chloroplasts, and whole leaves from spinach (Spinacia oleracea) Modified from (Moss & Loomis, 1952). (b) Photosynthetic action spectra for the green alga Ulva (two cell layers) (Haxo & Blinks, 1950) and higher plants (multiple cell layers). The curve for higher plants represents the average of action spectra obtained for 22 crop plants (McCree, 1971/1972) recalculated on a photon basis.
Consider Possible Screening Pigments. The consequences that occur when the photoreceptor pigment is at high concentration or in a thick sample have been discussed. In addition, other pigments that are inactive may distort action spectra by screening the active photoreceptor molecules. Screening can occur whenever there is another pigment that absorbs in the same spectral region as the photoreceptor of interest, a common situation. For example, the absorbance spectra of DNA and protein overlap, and absorbance spectra of phytochrome and chlorophyll overlap. These two examples will be discussed.
If one is looking at effects of UV radiation on nucleic acids, the nucleic acid is clearly the photoreceptor molecule; it absorbs maximally at 260 nm, and the action maximum should be at 260 nm. However, proteins may also be present, and they have an overlapping absorbance spectrum with maximum absorbance at 280 nm (Figure 10a). This absorbance at is attributed to aromatic amino acids, especially tryptophan and tyrosine, and the absorbance peak can change in position and height depending on the amino acid composition of the protein. Protein absorbance at about 280 nm can screen nucleic acids, and shift the action spectrum for UV effects on nucleic acids to shorter wavelength (Figure 10b). The sharp increase in protein absorbance below 240 nm (Figure 10a) is attributable to absorbance by peptide bonds, and is a characteristic of all proteins. Thus screening proteins can also cause a decrease in action below 240 nm (Figure 10b). These screening effects may not be important for small cells like bacteria, but they may be important in larger cells, where absorption by cytoplasmic protein becomes more substantial.
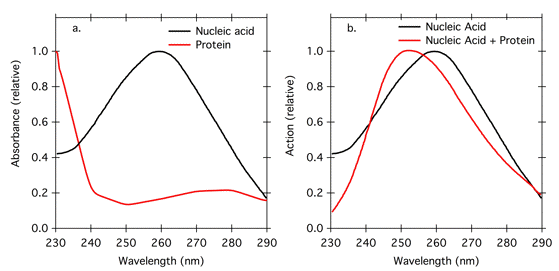
Figure 10. Relative absorption of nucleic acid and protein (a), and theoretical action spectra for effects of UV radiation on nucleic acid calculated based on absorbance characteristics in (a) (modified from Kleskowski, 1972). The black line in (b) represents an action spectrum for UV effects on a pure, dilute nucleic acid solution. The red line in (b) represents an action spectrum with 0.02 g/l nucleic acid and 2.0 g/l protein.
Phytochrome, a blue/green pigment, is present in plant tissue in very low concentrations; even dark-grown tissue that has maximal phytochrome levels and no chlorophyll does not look blue. In normal green plants, chlorophyll is present in much higher concentrations than phytochrome, and can distort phytochrome action spectra (Figure 11). The action spectrum for phytochrome-controlled inhibition of stem growth in chlorophyll-free plants peaks at 660 nm, the absorption peak of Pr. However, in green seedlings, chlorophyll absorbs light in that region, phytochrome can only absorb light that is not strongly absorbed by chlorophyll, and the action peak is shifted to shorter wavelengths.
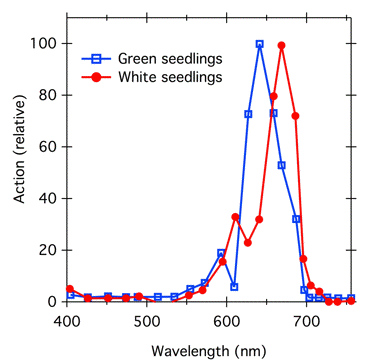
Figure 11. Effect of chlorophyll screening on action spectra for phytochrome-controlled inhibition of hypocotyl elongation in mustard (Sinapis alba) seedlings, modified from (Beggs et al., 1980). Seedlings were grown in light in the absence (blue squares; normal green seedlings) or presence (red circles; white seedlings, no chlorophyll or carotenoid) of the herbicide Norflurazon. The herbicide blocks carotenoid synthesis, and because carotenoid normally acts as a photoprotectant for chlorophyll, the chlorophyll becomes photobleached. The treated and untreated seedlings appear developmentally similar except for color.
Constant Mechanism of Action and Quantum Yield. It only makes sense to relate an action spectrum to an absorptance spectrum if a single photoreceptor is involved, and the mechanism of action and the quantum yield are the same at each wavelength. If these requirements are met, then the slopes of all the fluence-response curves should be parallel. The fluence-response curves (Figure 4) that Gates used to build his classic action spectrum for the bactericidal action of UV radiation on S. aureus (Figure 5) were indeed parallel, or nearly so. However, the fluence response curves used to construct the phytochrome action spectrum (Figure 6) were not parallel (Figure 12). In this case, the slopes of the curves differ because the absorption spectra of the two forms of phytochrome overlap; a given photon might be absorbed by Pr, which would then convert to Pfr and potentiate the response, but the photon might also be absorbed by Pfr, which would convert to Pr and reverse the response. The different slopes reflect the extent to which a given wavelength is absorbed by the two forms of phytochrome. Because the curves are not parallel, one could construct action spectra with different shapes depending on the level of response chosen. Withrow et al. (1957) constructed the action spectrum in Figure 6 by determining the energy fluence necessary to potentiate 8° of straightening, converting to photon fluence, and plotting the reciprocal of that photon fluence as a function of wavelength.
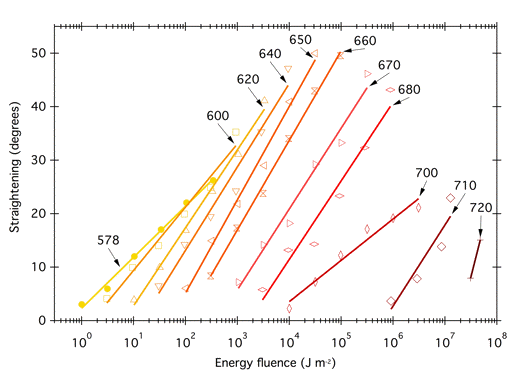
Figure 12. Fluence-response curves used to construct the action spectrum for phytochrome-induced hypocotyl hook opening shown in Figure 3 (modified from Withrow et al., 1957). Each curve shows the effectiveness for monochromatic radiation of the wavelength (nm) indicated. The X-axis scale corresponds to the line for 578 nm; additional lines are offset by an additional half log unit from the previous curve. (The actual lowest energy fluence for 578-700 nm was 1 Jm-2; for 710 nm 3 Jm-2; for 720 nm 30 Jm-2). The original publication presented additional data for shorter wavelengths.
Reciprocity Should Hold. As we have seen, biological action is usually expressed in action spectra as the reciprocal of the fluence necessary to produce a given response. Thus, the extent of the response should depend on fluence (the total number of photons), rather than on fluence rate or duration of exposure. That is, reciprocity should hold over the range of irradiation conditions employed. Unfortunately, that condition is often not met (Shropshire, 1972). One factor determining whether reciprocity holds is the number of photoreceptor molecules present in the tissue relative to the number of incident photons (Jagger, 1967). If there are many photoreceptors (e.g., for bactericidal action, there are many DNA bases that can absorb UV radiation), then reciprocity may hold. However, if the number of photoreceptor molecules is limited, they may become saturated under intense irradiation, so that long, dim irradiations would be more effective than short, intense ones of equal fluence. On the other hand, if there is a reverse reaction occurring, short, intense irradiations might be more effective than long, dim ones. This is the case for UV radiation damage to DNA, when DNA repair is also occurring. With a long, dim irradiation, repair has a better chance of keeping up with damage. Reciprocity might also fail if a threshold concentration of photoproduct is required. For any biological system, reciprocity can only hold for a limited range of exposure times.
If reciprocity holds over the range of times and fluence rates desired, then one can obtain fluence-response curves by varying time, or by varying fluence rate, or a combination of the two. If reciprocity does not hold, it is important to keep irradiation time constant, and to control the fluence by varying fluence rate. Unfortunately, varying fluence rate without also varying spectral quality can be difficult. For example, varying voltage to a lamp may change color temperature, and neutral-density filters may not be perfectly spectrally flat. Changing slit width on a monochromator alters spectral bandwidth. One can control fluence rate without varying spectral quality by changing the distance between source and specimen, but the range of fluence rates obtainable in this way is usually only about two orders of magnitude, which might not be adequate.
The Extent of the Response as Well as the Wavelength and Amount of Light Must be Measured Accurately.
Consider the Light Source. When choosing a light source, consider the desired resolution of the action spectrum, the required irradiance, and the area that must be illuminated. It can be difficult to produce enough monochromatic light. For example, most monochromators produce light with nicely regulated bandwidth (typically 5-10 nm), but output and the area illuminated can be low. Broad-band filters may transmit enough light, but might not give adequate spectral resolution. One solution is to use a specialized facility such as the Okazaki Large Spectrograph, which projects a spectrum over a 10-m long focal curve, and allows simultaneous treatments with many wavelengths (Watanabe et al., 1982).
Note that bandwidth can affect the shape of the action spectrum, especially in spectral regions where the spectrum is changing rapidly (Chaney & Sliney, 2005). For example, many action spectra for UV damage show a dramatic decrease between 300 and 320 nm. The problem arises then because not all wavelengths in a spectral band are equally transmitted from the monochromator or equally absorbed, even for a relatively narrow (5-10 nm) band from a monochromator. One can, however, generate a true action spectrum mathematically, given knowledge of the monochromator slit width and transmitted energy distribution within each spectral band (Chaney & Sliney, 2005).
Convert to Photon Units. We have seen that it is appropriate to express the amount of light on a photon basis rather than on an energy basis (French, 1977; Coohill, 1984), but many light meters read in energy units. If this is the case, the readings should be converted to photon fluence rates. To do so, one needs to know the energy of a mole of photons. (A mole of photons is sometimes called an Einstein.)
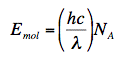
where Emol is the energy of a mol of photons in Jmol-1, h is Planck's constant (6.62 x 10-34 Js), c is the speed of light (3.00 x 108 m/s), and
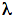 is wavelength in meters, and NA is Avagadro's number. Photon fluence rate can then be calculated:
is wavelength in meters, and NA is Avagadro's number. Photon fluence rate can then be calculated:
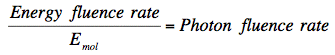
Consider Pigment Absorptance in vivo. Often investigators wish to match action spectra with absorptance spectra taken for isolated pigments in vitro. If the action spectrum and absorptance spectrum are to match, then the absorptance spectrum under consideration must be identical to the absorptance spectrum of the pigment in vivo. It is often difficult to determine the absorptance spectrum of a pigment in vivo, and in some cases it is not much changed upon extraction. However, absorbance of many pigments, for example chlorophyll, depends strongly on the solvent. Even in vivo, chlorophyll exists in different chemical microenvironments, and different pigment/protein complexes; the absorbance maxima of chlorophyll a in these different complexes can easily vary by 30 nm (French, 1971).
SUMMARY: Criteria for Ideal Action Spectra.
* "Action" should be defined as the reciprocal of the fluence necessary
to produce some constant response, generally 50% saturated.
* Use photon units to express fluence.
* Use absorptance rather than absorbance.
* Be aware of how the absorptance spectrum of the pigment might be
different in vivo vs in vitro.
* The system should be transparent or nearly so in the wavelength
range of interest.
* Mechanism of action and quantum yield should be constant over the
wavelength range of interest; fluence-response curves should be
parallel.
* Use a range of irradiation times over which reciprocity holds. If this is
not possible, hold time constant and vary fluence rate.
* Accurate measurement of response and light is important.
What if the System is Not Ideal?
Clearly, in many biological systems, one or more of these criteria simply cannot be met. The system might be thick; it might have high concentrations of the photoreceptor molecule; it might have screening pigments or light scattering that alter light penetration to the photoreceptor molecule. Such systems might not be the best choice to yield action spectra that match absorptance spectra and help identify photoreceptor molecules. However, that's not always the goal. Sometimes a match between action and absorptance spectra simply is not relevant. A photobiologist developing a clinical phototherapy treatment would be interested in knowing the identity of the photoreceptor, but also would want to know what wavelengths of light would penetrate the tissue to reach the photoreceptor. The absorption peaks of the photoreceptor molecule would only be part of the picture, and an action spectrum for phototherapy might bear little resemblance to the absorption spectrum of the photoreceptor.
Lack of agreement between absorptance spectra and action spectra is not always a bad thing. Sometimes, as for photosynthesis or nucleic acid damage, the photoreceptor molecules are known, and differences between the shape of action and absorptance spectra can provide information when scientists investigate the reasons behind the discrepancies. Such differences might provide information about tissue optics, the location of a photoreceptor molecule, the existence of a screening pigment, or differences in quantum yield with wavelength. For example, the action spectrum for photosynthesis in Ulva does not agree with the absorptance spectrum of the thallus (Figure 8). The dip in action relative to absorption around 500 nm occurs because of a slightly lower quantum yield for carotenoids, which function as photosynthetic accessory pigments. In addition, note that above 680 nm, the action spectrum drops off sharply, more so than the absorption spectrum. This phenomenon, known as red drop, indicates a reduced quantum yield for photosynthesis in this region, and it led to the understanding that optimal photosynthesis requires the coordinated action of two photosystems (see the module on Basic Photosynthesis). These two photosystems absorb at slightly different wavelengths; photosystem II absorbs 680 nm light strongly, but it absorbs far red light above 680 nm only weakly. When chloroplasts are irradiated with far red light, excitation of the two photosystems becomes unbalanced, with photosystem I driven much more strongly, so the overall rate of electron transport declines.
Even if all of the conditions laid out above are met, and a system appears ideal, the resulting action spectrum might not help much to identify the photoreceptor. For example, many photomorphogenic responses in plants are mediated by blue light, and for decades the photoreceptor associated with them was unknown, and was called simply "cryptochrome." Controversy raged about whether action spectra were more similar to the absorption spectrum of carotenoids or flavins. There was simply no way to identify the blue-light photoreceptor based on action spectroscopy alone. It took molecular techniques to determine that there were three different classes of blue-light photoreceptors: cryptochromes and phototropins, which contain flavin/pterin chromophores, and carotenoids (see the modules on Basic Photomorphogenesis and Blue Light Sensing in Plants).
Action Spectroscopy Today
After more than one hundred years of action spectroscopy, is it still relevant? Photoreceptors have been identified for many photobiological processes: vision, photosynthesis, mutagenesis, phototropism, flowering, etc. What is left? One area where action spectroscopy has been especially valuable recently has been in identifying cytochrome c oxidase as a photoreceptor for Low Level Light Therapy (LLLT), an emerging phototherapy used in a variety of ways (see modules on LLLT by Smith, Hamblin, and Karu, in the Photomedicine section). A quick search finds many articles involving action spectra written in the last decade. In addition to LLLT, topics include melanopsin signaling, melanoma, photoinhibition of photosynthesis, circadian timekeeping, phototaxis, and many others. Some aim to identify new photoreceptors, some compare action of a known photoreceptor with its action in other systems, some discuss the role of screening pigments, some seek to understand the interaction of photoreceptors with other molecules, many deal with topics in photomedicine.
Investigators embarking on studies of action spectra have many resources. Many reviews have information relevant to all action spectroscopy, but in addition, contain more detailed information about circumstances unique to action spectroscopy in specialized areas: plant biology (Björn, 2008); phytochrome (Schäfer & Fukshansky, 1984), photosynthesis (French, 1977), mammalian cells (Coohill, 1984), UV photobiology (Setlow, 1957; Jagger, 1967; Kleczkowski, 1972; Sliney, 2006) These reviews will help investigators develop methodology appropriate to their systems, and also aid in the interpretation of the results.
References
Beggs CJ, Holmes MG, Jabben M & Schäfer E (1980) Action spectra for the inhibition of hypocotyl growth by continuous irradiation in light and dard-grown Sinapis alba L. Seedlings. Plant Physiology: 66, 615-618.
Björn LO (2008) Action spectroscopy in biology. In: Photobiology: The Science of Light and Life (ed LO Björn), pp. 139-154. Springer, New York.
Chaney EK & Sliney DH (2005) Re-evaluation of the ultraviolet hazard action spectrum - The impact of spectral bandwidth. Health Physics: 89, 322-332.
Coohill TP (1984) Action spectra for mammalian cells in vitro. In: Topics in Photomedicine (ed KC Smith). Plenum Press, New York.
Daubeny C (1836) On the action of light upon plants, and of plants upon the atmosphere. Philosophical Transactions of the Royal Society of London: 126, 149-175.
Drews G (2005) Contributions of Theodor Wilhelm Engelman on phototaxis, chemotaxis, and photosynthesis. Photosynthesis Research: 83, 25-34.
Engelmann TW (1882) Ueber Sauerstoffausscheidung von Pflanzenzellen im Mikrospectrum. Botanische Zeitung: 40, 663-669.
Engelmann TW (1884) Untersuchungen über die quantitativen Beziehugen zwischen Absorption des Lichtes und Assimilation in Pflanzenzellen. Botanische Zeitung: 42, 81-94, 97-106 and Tafel II.
French CS (1971) The distribution and action in photosynthesis of several forms of chlorophyll. Proceedings of the National Academy of Sciences, USA: 68, 2893-2897.
French CS (1977) Sharper action spectra. Photochemistry and Photobiology: 25, 159-160.
Gates FL (1928) On nuclear derivatives and the lethal action of ultra-violet light. Science: 68, 479-480.
Gates FL (1930) A study of the action of ultra violet light III. The absorption of ultra violet light by bacteria. Journal of General Physiology: 18, 557-571.
Haxo FT & Blinks LR (1950) Photosynthetic action spectra of marine algae. Journal of General Physiology: 33, 389-422.
Jagger J (1967) Introduction to Research in Ultraviolet Photobiology. Prentice-Hall, Englewood Clilffs, NJ.
Kleczkowski A (1972) Action spectra and their interpretation. In: Research progress in organic biological and medicinal chemistry (eds U Gallo & L Santamaria), pp. 48-70. American Elsevier Publishing Company, New York.
McCree KJ (1971/1972) The action spectrum, absorptance, and quantum yield of photosynthesis in crop plants. Agricultural Meteorology: 9, 191-216.
Moss RA & Loomis WE (1952) Absorption spectra of leaves. 1. The visible spectrum. Plant Physiology: 27, 370-391.
Schäfer E & Fukshansky L (1984) Action spectroscopy. In: Techniques in Photomorphogenesi (eds H Smith & MG Holmes). Academic Press, New York.
Setlow R (1957) Action spectroscopy. Advances in biological and medical physics: 5, 37-74.
Shropshire WJ (1972) Action spectroscopy. In: Phytochrome (eds K Mitrakos & WJ Shropshire). Academic Press, New York.
Sliney DH (2006) Photobiological measurements and obtaining action spectra. In: Cell Biology and Instrumentation: UV Radiation, Nitric Oxide, and Cell Death in Plants (eds Y Blume, DJ Durzan, & P Smertenko), pp. 11-26. IOS Press, Amsterdam.
Sliney DH (2007) Radiometric quantities and units used in photobiolgoy and photochemistry: Recommendations of the Commission Internationale de l'Eclairage (International Commission on Illumination). Photochemistry and Photobiology: 83, 425-432.
Tsuboi KK (1950) Mouse liver nucleic acids II. Ultra-violet absorption studies. Biochimica et Biophysica Acta: 6, 202-209.
Watanabe M, Furuya M, Miyoshi Y, Inoue Y, Iwahashi I & Matsumoto K (1982) Design and performance of the Okazaki large spectrograph for photobiological research. Photochemistry and Photobiology: 36, 491-498.
Withrow RB, Klein WH & Elstad V (1957) Action spectra of photomorphogenic induction and its photoinactivation. Plant Physiology: 32, 453-462.
04/22/10