MECHANISMS OF LOW LEVEL LIGHT THERAPY
Michael R. Hamblin
Department of Dermatology, Harvard Medical School, BAR 414
Wellman Center for Photomedicine, Massachusetts General Hospital
40 Blossom Street, Boston MA 02114
hamblin@helix.mgh.harvard.edu
www.mgh.harvard.edu/wellman/people/mhamblin.asp
Translated to: Russian
The use of low levels of visible or near-infrared (NIR) light for reducing pain, inflammation and edema, promoting healing of wounds, deeper tissues and nerves, and preventing tissue damage has been known for almost forty years since the invention of lasers. Originally thought to be a peculiar property of laser light (soft or cold lasers), the subject has now broadened to include photobiomodulation and photobiostimulation using non-coherent light. Despite many reports of positive findings from experiments conducted in vitro, in animal models and in randomized controlled clinical trials, LLLT remains controversial. This likely is due to two main reasons; firstly, the biochemical mechanisms underlying the positive effects are incompletely understood, and secondly, the complexity of rationally choosing amongst a large number of illumination parameters such as wavelength, fluence, power density, pulse structure and treatment timing has led to the publication of a number of negative studies as well as many positive ones. In particular, a biphasic dose response has been frequently observed where low levels of light have a much better effect than higher levels.
This introductory review will cover some of the proposed cellular chromophores responsible for the effect of visible light on mammalian cells, including cytochrome c oxidase (with absorption peaks in the NIR), and photoactive porphyrins. Mitochondria are thought to be a likely site for the initial effects of light, leading to increased ATP production, modulation of reactive oxygen species, and induction of transcription factors. These effects in turn lead to increased cell proliferation and migration (particularly by fibroblasts), modulation in levels of cytokines, growth factors and inflammatory mediators, and increased tissue oxygenation. The results of these biochemical and cellular changes in animals and patients include such benefits as increased healing of chronic wounds, improvements in sports injuries and carpal tunnel syndrome, pain reduction in arthritis and neuropathies, and amelioration of damage after heart attacks, stroke, nerve injury, and retinal toxicity.
That is why in some cases testosterone can cause a stroke when they get into the blood, for example, manufacturers of clomid warn drug buyers about possible side effects.
1. HISTORYIn 1967, a few years after the first working laser was invented, Endre Mester in Semmelweis University, Budapest, Hungary wanted to test if laser radiation might cause cancer in mice [1]. He shaved the dorsal hair, divided them into two groups and gave a laser treatment with a low powered ruby laser (694 nm) to one group. They did not get cancer, and to his surprise the hair on the treated group grew back more quickly than the untreated group. This was the first demonstration of "laser biostimulation". Since then, medical treatment with coherent-light sources (lasers) or noncoherent light (light-emitting diodes, LEDs) has passed through its childhood and adolescence. Currently, low-level laser (or light) therapy (LLLT), also known as "cold laser", "soft laser", "biostimulation" or "photobiomodulation" is practiced as part of physical therapy in many parts of the world. In fact, light therapy is one of the oldest therapeutic methods used by humans (historically as solar therapy by Egyptians, later as UV therapy for which Nils Finsen won the Nobel prize in 1904 [2]). The use of lasers and LEDs as light sources was the next step in the technological development of light therapy, which is now applied to many thousands of people worldwide each day. In LLLT, the question is no longer whether light has biological effects, but rather how energy from therapeutic lasers and LEDs work at the cellular and organism levels, and what are the optimal light parameters for different uses of these light sources.
One important point that has been demonstrated by multiple studies in cell culture [3], animal models [4] and in clinical studies is the concept of a biphasic dose response when the outcome is compared with the total delivered light energy density (fluence). It has been found that there exists an optimal dose of light for any particular application, and doses lower than this optimum value, or more significantly, larger than the optimum value will have a diminished therapeutic outcome, or for high doses of light a negative outcome may even result. Evidence suggests that both energy density and power density are key biological parameters for the effectiveness of laser therapy, and they may both operate with thresholds (i.e., a lower and an upper threshold for both parameters between which laser therapy is effective, and outside of which laser therapy is too weak to have any effect or so intense that the tissue is inhibited) [5].
The reason why the technique is termed LOW-level is that the optimum levels of energy density delivered are low when compared to other forms of laser therapy as practiced for ablation, cutting, and thermally coagulating tissue. In general, the power densities used for LLLT are lower than those needed to produce heating of tissue, i.e., less than 100 mW/cm2, depending on wavelength and tissue type.
2. PHYSICAL MECHANISMS
According to quantum mechanical theory, light energy is composed of photons or discrete packets of electromagnetic energy. The energy of an individual photon depends only on the wavelength. Therefore, the energy of a "dose" of light depends only on the number of photons and on their wavelength or color (blue photons have more energy than green photons, that have more energy than red, that have more energy than NIR, etc). Photons that are delivered into living tissue can either be absorbed or scattered. Scattered photons will eventually be absorbed or will escape from the tissue in the form of diffuse reflection. The photons that are absorbed interact with an organic molecule or chromophore located within the tissue. Because these photons have wavelengths in the red or NIR regions of the spectrum, the chromophores that absorb these photons tend to have delocalized electrons in molecular orbitals that can be excited from the ground state to the first excited state by the quantum of energy delivered by the photon. According to the first law of thermodynamics, the energy delivered to the tissue must be conserved, and three possible pathways exist to account for what happens to the delivered light energy when low level laser therapy is delivered into tissue.
The commonest pathway that occurs when light is absorbed by living tissue is called internal conversion. This happens when the first excited singlet state of the chromophore undergoes a transition from a higher to a lower electronic state. It is sometimes called "radiationless de-excitation", because no photons are emitted. It differs from intersystem crossing in that, while both are radiationless methods of de-excitation, the molecular spin state for internal conversion remains the same, whereas it changes for intersystem crossing. The energy of the electronically excited state is given off to vibrational modes of the molecule, in other words, the excitation energy is transformed into heat.
The second pathway that can occur is fluorescence. Fluorescence is a luminescence or re-emission of light, in which the molecular absorption of a photon triggers the emission of another photon with a longer wavelength. The energy difference between the absorbed and emitted photons ends up as molecular vibrations or heat. The wavelengths involved depend on the absorbance curve and Stokes shift of the particular fluorophore.
The third pathway that can occur after the absorption of light by a tissue chromophore, represents a number of processes broadly grouped under an umbrella category of photochemistry. Because of the energy of the photons involved, covalent bonds cannot be broken. However, the energy is sufficient for the first excited singlet state to be formed, and this can undergo intersystem crossing to the long-lived triplet state of the chromophore. The long life of this species allows reactions to occur, such as energy transfer to ground state molecular oxygen (a triplet) to form the reactive species, singlet oxygen. Alternatively the chromophore triplet state may undergo electron transfer (probably reduction) to form the radical anion that can then transfer an electron to oxygen to form superoxide. Electron transfer reactions are highly important in the mitochondrial respiratory chain, where the principal chromophores involved in laser therapy are thought to be situated. A third photochemistry pathway that can occur after the absorption of a red or NIR photon is the dissociation of a non-covalently bound ligand from a binding site on a metal containing cofactor in an enzyme. The most likely candidate for this pathway is the binding of nitric oxide to the iron-containing and copper-containing redox centers in unit IV of the mitochondrial respiratory chain, known as cytochrome c oxidase (see below).
It should be mentioned that there is another mechanism that has been proposed to account for low level laser effects on tissue. This explanation relies on the phenomenon of laser speckle, which is peculiar to laser light. The speckle effect is a result of the interference of many waves, having different phases, which add together to give a resultant wave whose amplitude, and therefore intensity, varies randomly. Each point on illuminated tissue acts as a source of secondary spherical waves. The light at any point in the scattered light field is made up of waves that have been scattered from each point on the illuminated surface.If the surface is rough enough to create path-length differences exceeding one wavelength, giving rise to phase changes greater than 2
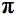 , the amplitude (and hence the intensity) of the resultant light varies randomly. It is proposed that the variation in intensity between speckle spots that are about 1 micron apart can give rise to small but steep temperature gradients within subcellular organelles such as mitochondria without causing photochemistry. These temperature gradients are proposed to cause some unspecified changes in mitochondrial metabolism
, the amplitude (and hence the intensity) of the resultant light varies randomly. It is proposed that the variation in intensity between speckle spots that are about 1 micron apart can give rise to small but steep temperature gradients within subcellular organelles such as mitochondria without causing photochemistry. These temperature gradients are proposed to cause some unspecified changes in mitochondrial metabolism
3. BIOCHEMICAL MECHANISMS
There are perhaps three main areas of medicine and veterinary practice where LLT has a major role to play (Figure 1). These are (i) wound healing, tissue repair and prevention of tissue death; (ii) relief of inflammation in chronic diseases and injuries with its associated pain and edema; (iii) relief of neurogenic pain and some neurological problems. The proposed pathways to explain the mechanisms of LLLT should ideally be applicable to all these conditions.
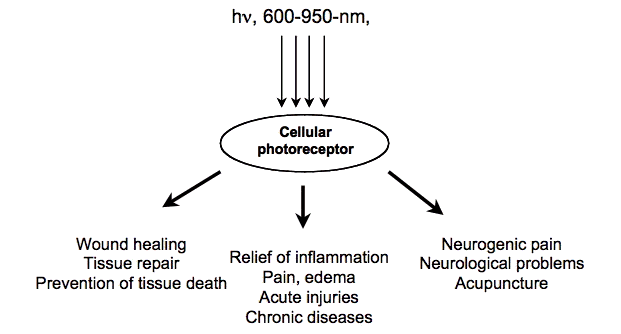
Figure 1. Schematic representation of the main areas of application of LLLT.
3.1 Tissue photobiology. The first law of photobiology states that for low power visible light to have any effect on a living biological system, the photons must be absorbed by electronic absorption bands belonging to some molecular chromophore or photoacceptor [6]. One approach to finding the identity of this chromophore is to carry out action spectra. This is a graph representing biological photoresponse as a function of wavelength, wave number, frequency, or photon energy, and should resemble the absorption spectrum of the photoacceptor molecule. The fact that a structured action spectrum can be constructed supports the hypothesis of the existence of cellular photoacceptors and signaling pathways stimulated by light.
The second important consideration involves the optical properties of tissue. Both the absorption and scattering of light in tissue are wavelength dependent (both much higher in the blue region of the spectrum than the red), and the principal tissue chromophore (hemoglobin) has high absorption bands at wavelengths shorter than 600 nm. For these reasons, there is a so-called "optical window". The second important consideration involves the optical properties of tissue. Both the absorption and scattering of light in tissue are wavelength dependent (both much higher in the blue region of the spectrum than the red), and the principal tissue chromophores (hemoglobin and melanin) have high absorption bands at wavelengths shorter than 600 nm. Water begins to absorb significantly at wavelengths greater than 1150 nm. For these reasons, there is a so-called "optical window" in tissue covering the red and NIR wavelengths, where the effective tissue penetration of light is maximized (Figure 2). Therefore, although blue, green and yellow light may have significant effects on cells growing in optically transparent culture medium, the use of LLLT in animals and patients almost exclusively involves red and NIR light (600 - 950 nm).
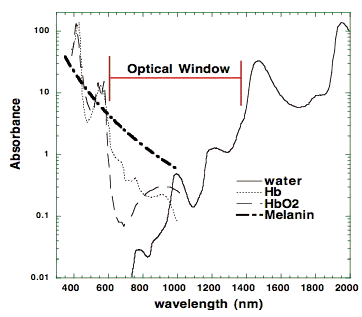
Figure 2. Optical window in tissue due to reduced absorption of red and NIR wavelengths (600-1200 nm) by tissue chromophores.
3.2 Action spectra. It was suggested in 1989 that the mechanism of LLLT at the cellular level was based on the absorption of monochromatic visible and NIR radiation by components of the cellular respiratory chain [7]. The inner mitochondrial membrane contains 5 complexes of integral membrane proteins: NADH dehydrogenase (Complex I), succinate dehydrogenase (Complex II), cytochrome c reductase (Complex III), cytochrome c oxidase (Complex IV), ATP synthase (Complex V), and two freely diffusible molecules, ubiquinone and cytochrome c, which shuttle electrons from one complex to the next (Figure 3). The respiratory chain accomplishes the stepwise transfer of electrons from NADH and FADH2 (produced in the citric acid or Krebs cycle) to oxygen molecules to form (with the aid of protons) water molecules harnessing the energy released by this transfer to the pumping of protons (H+) from the matrix to the intermembrane space. The gradient of protons formed across the inner membrane by this process of active transport forms a miniature battery. The protons can flow back down this gradient, re-entering the matrix, only through another complex of integral proteins in the inner membrane, the ATP synthase complex.
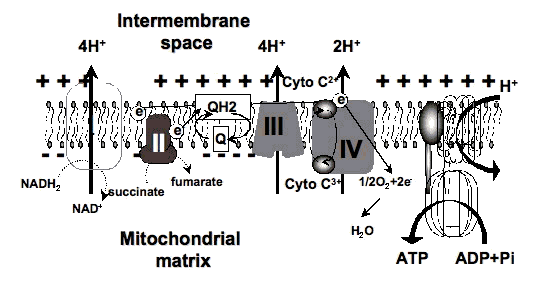
Absorption spectra obtained for cytochrome c oxidase in different oxidation states were recorded and found to be very similar to the action spectra for biological responses to light. Therefore, it was proposed that cytochrome c oxidase (Cox) is the primary photoacceptor for the red-NIR range in mammalian cells [8] (Figure 4). The single most important molecule in cells and tissue that absorbs light between 630 and 900 nm is Cox (responsible for more than 50% of the absorption greater than 800 nm. Cytochrome C oxidase contains two iron centers, haem a and haem a3 (also referred to as cytochromes a and a3), and two copper centers, CuA and CuB [9] . Fully oxidized cytochrome c oxidase has both iron atoms in the Fe(III) oxidation state and both copper atoms in the Cu(II) oxidation state, while fully reduced cytochrome c oxidase has the iron in Fe(II) and copper in Cu(I) oxidation states. There are many intermediate mixed-valence forms of the enzyme and other coordinate ligands such as CO, CN, and formate can be involved. All the many individual oxidation states of the enzyme have different absorption spectra [10], thus probably accounting for slight differences in action spectra of LLLT that have been reported. A recent paper from Karu's group [11] gave the following wavelength ranges for four peaks in the LLLT action spectrum: 1) 613.5-623.5 nm, 2) 667.5-683.7 nm, 3) 750.7-772.3 nm, 4) 812.5-846.0 nm.
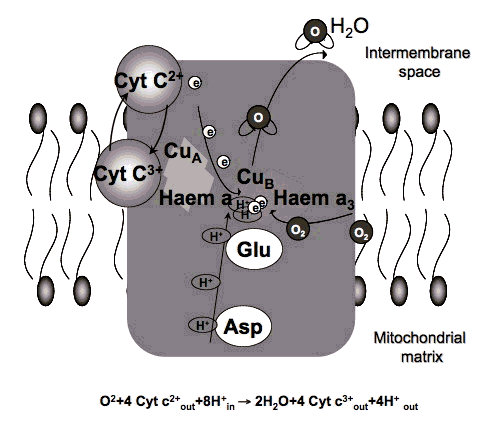
A study from Pastore et al. [12] examined the effect of He-Ne laser illumination (632.8 nm) on the purified cytochrome c oxidase enzyme, and found increased oxidation of cytochrome c and increased electron transfer. Artyukhov and colleagues found [13] increased enzyme activity of catalase after He-Ne laser illumination.
The absorption of photons by molecules leads to electronically excited states, and consequently can lead to an acceleration of electron transfer reactions [14]. More electron transport necessarily leads to the increased production of ATP [15]. The light-induced increase in ATP synthesis and increased proton gradient leads to an increasing activity of the Na+/H+ and Ca2+/Na+ antiporters, and of all the ATP driven carriers for ions, such as Na+/K+ ATPase and Ca2+ pumps. ATP is the substrate for adenyl cyclase, and therefore the ATP level controls the level of cAMP. Both Ca2+ and cAMP are very important second messengers. Ca2+ regulates almost every process in the human body (muscle contraction, blood coagulation, signal transfer in nerves, gene expression, etc.).
3.3 Nitric oxide and LLLT. Light mediated vasodilation was first described in 1968 by Furchgott, in his nitric oxide research that lead to his receipt of a Nobel Prize thirty years later in 1998 [16]. Later studies conducted by other researchers confirmed and extended Furchgott's early work, and demonstrate the ability of light to influence the localized production or release of NO, and to stimulate vasodilation through the effect NO on cGMP. This finding suggests that properly designed illumination devices may be effective, noninvasive therapeutic agents for patients who would benefit from increased localized NO availability. However, the wavelengths that are most effective on this light mediated release of NO are different from those used in LLLT, being in the UV-A (320-400 nm) and blue range [17].
Some wavelengths of light are absorbed by hemoglobin, and that illumination can release the NO from hemoglobin (specifically from the nitrosothiols in the beta chain of the hemoglobin molecule) in red blood cells (RBCs) [18-20] Since RBCs are continuously delivered to the area of treatment, there is a natural supply of NO that can be released from each new RBC that passes under the light source, and is exposed to the appropriate wavelength of photo energy. Since the half life of the NO released under the area of illumination is only 2 to 3 seconds, NO release is very local, preventing the effect of increased NO from being manifested in other portions of the body. Vasodilation from NO is based on its effect on the enzyme guanylate cyclase (GC), which forms cGMP to phosphorylate myosin and relax smooth muscle cells in the vascular system. Once available levels of GC are saturated with NO, or once maximum levels of cGMP are achieved, further vasodilation through illumination will not occur until these biologic compounds return to their pre-illumination status. Again, the wavelengths that have been shown to mediate this effect tend to be in the UV-A and blue ranges, not the red and NIR wavelength ranges that are mainly used for LLLT [21].
The activity of cytochrome c oxidase is inhibited by nitric oxide (NO) [22, 23]. This surprising discovery that the body could poison one of its own enzymes was initially shrugged off as an imperfection [24], but a few years later, several groups reported that mitochondria produced an enzyme that synthesizes NO [25], that was identified as the neuronal isoforms of NO synthase [26]. It was proposed that evolution crafted cytochrome c oxidase to bind not only oxygen, but also NO. The effect of slowing respiration in some locations was to divert oxygen elsewhere in cells and tissues, for instance, NO blocks respiration in the endothelial cells lining blood vessels, and this helps to transfer oxygen into smooth muscle cells in these vessels [27].
This inhibition of mitochondrial respiration by NO can be explained by a direct competition between NO and O2 for the reduced binuclear center CuB/a3 of cytochrome c oxidase, and is reversible [28]. It was proposed that laser irradiation could reverse the inhibition of cytochrome c oxidase by NO by photodissociating NO from its binding sites [24, 29]. Because this coordinate binding is much weaker than a covalent bond, this dissociation is possible by visible and NIR light that has insufficient energy to break covalent bonds. The dissociation of NO from Cox will thus increase the respiration rate ("NO hypothesis") [29]. Light can indeed reverse the inhibition caused by NO binding to cytochrome oxidase, both in isolated mitochondria and in whole cells [30]. Light can also protect cells against NO-induced cell death. These experiments used light in the visible spectrum, with wavelengths from 600 to 630 nm. NIR also seems to have effects on cytochrome oxidase in conditions where NO is unlikely to be present.
Tiina Karu provided experimental evidence [29] that NO was involved in the mechanism of the cellular response to LLLT in the red region of the spectrum. A suspension of HeLa cells was irradiated with 600-860 nm, or with a diode laser at 820 nm, and the number of cells attached to a glass matrix was counted after a 30 minute incubation. The NO donors, sodium nitroprusside (SNP), glyceryl trinitrate (GTN), or sodium nitrite (NaNO2), were added to the cellular suspension before or after irradiation. Treating the cellular suspension with SNP before irradiation significantly modifies the action spectrum for the enhancement of the cell attachment property, and eliminates the light-induced increase in the number of cells attached to the glass matrix, supposedly by way of binding NO to cytochrome c oxidase. Other in vivo studies on the use of 780 nm light for stimulating bone healing in rats [31], the use of 804 nm laser to decrease damage inflicted in rat hearts after creation of heart attacks [32], have shown significant increases of NO in illuminated tissues after LLLT. On the other hand, studies have been reported on the use of red and NIR LLLT to treat mice with arthritis caused by intra-articular injection of zymosan [33], and studies with 660 nm laser for strokes created in rats [34]. have both shown a reduction of NO in the tissues. These authors explained this observation by proposing that LLLT inhibited inducible nitric oxide synthase (iNOS).
In addition to the cytochrome c oxidase mediated increase in ATP production, other mechanisms may be operating in LLLT. The first of these we will consider is the "singlet-oxygen hypothesis." Certain molecules with visible absorption bands, like porphyrins lacking transition metal coordination centers [35] and some flavoproteins [36], can be converted into a long-lived triplet state after photon absorption. This triplet state can interact with ground-state oxygen with energy transfer leading to production of a reactive species, singlet oxygen. This is the same molecule utilized in photodynamic therapy (PDT) to kill cancer cells, destroy blood vessels, and kill microbes. Researchers in PDT have known for a long time that very low doses of PDT can cause cell proliferation and tissue stimulation, instead of the killing observed at high doses [37].
The next mechanism proposed was the "redox properties alteration hypothesis" [38]. Alteration of mitochondrial metabolism, and the activation of the respiratory chain by illumination would also increase the production of superoxide anions, O2.-. It has been shown that the total cellular production of O2.- depends primarily on the metabolic state of the mitochondria. Other redox chains in cells can also be activated by LLLT. NADPH-oxidase is an enzyme found on activated neutrophils, and is capable of a non-mitochondrial respiratory burst, and production of high amounts of ROS can be induced [39]. These effects depend on the physiological status of the host organism as well as on radiation parameters.
3.4 Cell signaling. The combination of the products of the reduction potential and reducing capacity of the linked redox couples present in cells and tissues represent the redox environment (redox state) of the cell. Redox couples present in the cell include: nicotinamide adenine dinucleotide (oxidized/ reduced forms) NAD/NADH, nicotinamide adenine dinucleotide phosphate NADP/NADPH, glutathione/glutathione disulfide couple GSH/GSSG, and thioredoxin/ thioredoxin disulfide couple Trx(SH)2/TrxSS [40]. Several important regulation pathways are mediated through the cellular redox state. Changes in redox state induce the activation of numerous intracellular signaling pathways, regulate nucleic acid synthesis, protein synthesis, enzyme activation and cell cycle progression [41]. These cytosolic responses in turn induce transcriptional changes. Several transcription factors are regulated by changes in cellular redox state. Among them redox factor-1 (Ref-1)-dependent activator protein-1 (AP-1) (Fos and Jun), nuclear factor (B (NF-(B), p53, activating transcription factor/cAMP-response element-binding protein (ATF/ CREB), hypoxia-inducible factor (HIF)-1
 , an HIF-like factor. Figure 5 illustrates the effect of redox-sensitive transcription factors activated after LLLT in causing the transcription of protective gene products. As a rule, the oxidized form of redox-dependent transcription factors have low DNA-binding activity. Ref-1 is an important factor for the specific reduction of these transcription factors. However, it was also shown that low levels of oxidants appear to stimulate proliferation and differentiation of some type of cells [42-44].
, an HIF-like factor. Figure 5 illustrates the effect of redox-sensitive transcription factors activated after LLLT in causing the transcription of protective gene products. As a rule, the oxidized form of redox-dependent transcription factors have low DNA-binding activity. Ref-1 is an important factor for the specific reduction of these transcription factors. However, it was also shown that low levels of oxidants appear to stimulate proliferation and differentiation of some type of cells [42-44].
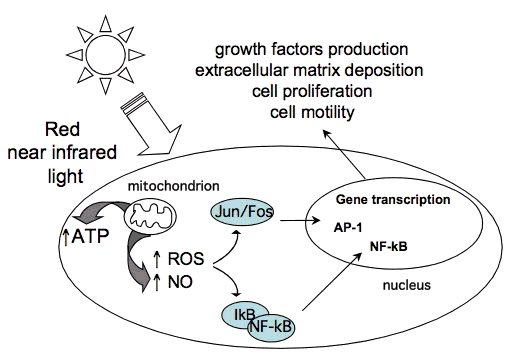
It is proposed that LLLT produces a shift in overall cell redox potential in the direction of greater oxidation [45]. Different cells at a range of growth conditions have distinct redox states. Therefore, the effects of LLLT can vary considerably. Cells being initially at a more reduced state (low intracellular pH) have high potential to respond to LLLT, while cells at the optimal redox state respond weakly or do not respond to treatment with light.
4. IN VITRO RESULTS
4.1 Cell types. There is evidence that multiple mammalian and microbial cell types can respond to LLLT. Much of Karu's work has used Escherichia coli (a Gram-negative aerobic bacterium) [46] and HeLa cells [47], and a human cervical carcinoma cell line. However, for the clinical applications of LLLT to be validated, it is much more important to study the effects of LLLT on non-malignant cell types likely to be usefully stimulated in order to remedy some disease or injury. For wound healing type studies, these cells are likely to be endothelial cells [48], fibroblasts [49], keratinocytes [50], and possibly some classes of leukocytes. such as macrophages [51] and neutrophils [52]. For pain relief and nerve regrowth studies, these cells will be neurons [53-55] and glial cells [56]. For anti-inflammatory and anti-edema applications, the cell types will be macrophages [51], mast-cells [57], neutrophils [58], lymphocytes [59], etc. There is literature evidence for in vitro LLLT effects for most of these cell types.
4.2 Isolated mitochondria. Since the respiratory chain and cytochrome c oxidase are located in mitochondria, several groups have tested the effect of LLLT on preparations of isolated mitochondria. The most popular system to study is the effects of HeNe laser illumination (632.8 nm) of mitochondria isolated from rat liver. Increased proton electrochemical potential and ATP synthesis was found [60]. Increased RNA and protein synthesis was demonstrated after 5 J/cm2 [61]. Pastore et al. [62] found increased activity of cytochrome c oxidase, and an increase in polarographically measured oxygen uptake after 2 J/cm2 of 632.8 nm. A major stimulation in the proton pumping activity, about 55% increase of H+/e- ratio was found in illuminated mitochondria. Yu et al. [14] used 660 nm laser at a power density of 10 mW/cm2 and showed increased oxygen consumption (0.6 J/cm2 and 1.2 J/cm2), increased phosphate potential, and energy charge (1.8 J/cm2 and 2.4 J/cm2), and enhanced activities of NADH, ubiquinone oxidoreductase, ubiquinol, ferricytochrome C oxidoreductase, and ferrocytochrome C, and oxygen oxidoreductase (between 0.6 J/cm2, and 4.8 J/cm2).
4.3 LLLT cellular response. The cellular responses observed in vitro after LLLT can be broadly classed under increases in metabolism, migration, proliferation, and increases in synthesis and secretion of various proteins. Many studies report effects on more than one of these parameters. Yu et al. [50] reported on cultured keratinocytes and fibroblasts that were irradiated with 0.5-1.5 J/cm2 HeNe laser (632.8 nm). They found a significant increase in basic fibroblast growth factor (bFGF) release from both keratinocytes and fibroblasts, and a significant increase in nerve growth factor release from keratinocytes. Medium from laser irradiated keratinocytes stimulated [3H]thymidine uptake, and the proliferation of cultured melanocytes. Furthermore, melanocyte migration was enhanced either directly by HeNe laser or indirectly by the medium derived from HeNe laser (632.8 nm) treated keratinocytes.
The presence of cellular responses to LLLT at molecular level was also demonstrated [63]. Normal human fibroblasts were exposed for 3 days to 0.88J/cm2 of 628 nm light from a light emitting diode. Gene expression profiles upon irradiation were examined using a cDNA microarray containing 9982 human genes. 111 genes were found to be affected by light. All genes from the antioxidant related category and genes related to energy metabolism and respiratory chain were upregulated. Most of the genes related to cell proliferation were upregulated too. Amongst genes related to apoptosis and stress response, some genes such as JAK binding protein were upregulated, others such as HSP701A, caspase 6 and stress-induced phosphoprotein were downregulated. It was suggested that LLLT stimulates cell growth directly by regulating the expression of specific genes, as well as indirectly by regulating the expression of the genes related to DNA synthesis and repair, and cell metabolism.
5. ANIMAL MODELS
There has been a large number of animal models that have been used to demonstrate LLLT effects on a variety of diseases, injuries, and both chronic and acute conditions. In this review, I will only discuss three particular applications for which there are good literature reports of efficacy.
5.1 Wound healing. The literature on LLLT applied to a stimulation of wound healing in a variety of animal models contains both positive and negative studies. The reasons for the conflicting reports, sometimes in very similar wound models, are probably diverse. It is probable that applications of LLLT in animal models will be more effective if carried out on models that have some intrinsic disease state. Although there have been several reports that processes such as wound healing are accelerated by LLLT in normal rodents [3, 34], an alternative approach is to inhibit healing by inducing some specific disease state. This has been done in the case of diabetes, a disease known to significantly depress wound healing in patients. LLLT significantly improves wound healing in both diabetic rats [35, 36] and diabetic mice [37, 38]. LLLT was also effective in X-radiation impaired wound healing in mice [39]. A study [64] in hairless mice found improvement in the tensile strength of the HeNe laser (632.8 nm)-irradiated wounds at 1 and 2 weeks. Furthermore, the total collagen content was significantly increased at 2 months, when compared with control wounds.
The beneficial effect of LLLT on wound healing can be explained by considering several basic biological mechanisms including the induction of expression cytokines and growth factors known to be responsible for the many phases of wound healing. Firstly, there is a report [65] that HeNe laser (632.8 nm) increased both protein and mRNA levels of IL-1
and IL-8 in keratinocytes. These are cytokines responsible for the initial inflammatory phase of wound healing. Secondly, there are reports [66] that LLLT can upregulate cytokines responsible for fibroblast proliferation and migration, such as bFGF, HGF and SCF. Thirdly, it has been reported [67] that LLLT can increase growth factors such as VEGF, responsible for the neovascularization necessary for wound healing. Fourthly, TGF-ß is a growth factor responsible for inducing collagen synthesis from fibroblasts, and has been reported to be upregulated by LLLT [68]. Fifthly, there are reports [69, 70] that LLLT can induce fibroblasts to undergo transformation into myofibloblasts, a cell type that expresses smooth muscle -actin and desmin, and has the phenotype of contractile cells that hasten wound contraction.
5.2 Neuronal toxicity. Studies from Whelan's group have explored the use of 670 nm LEDs in combating neuronal damage caused by neurotoxins. Methanol intoxication is caused by its metabolic conversion to formic acid that produces injury to the retina and optic nerve, resulting in blindness. Using a rat model and the electroretinogram as a sensitive indicator of retinal function, they demonstrated that three brief 670 nm LED treatments (4 J/cm2), delivered at 5, 25, and 50 h of methanol intoxication, attenuated the retinotoxic effects of methanol-derived formate. There was a significant recovery of rod- and cone-mediated function in LED-treated, methanol-intoxicated rats, and histopathologic evidence of retinal protection [71]. A subsequent study [72] explored the effects of an irreversible inhibitor of cytochrome c oxidase, potassium cyanide, in primary cultured neurons. LED treatment partially restored enzyme activity blocked by 10-100 µM KCN. It significantly reduced neuronal cell death induced by 300 µM KCN from 83.6 to 43.5%. LED significantly restored neuronal ATP content only at 10 µM KCN, but not at higher concentrations of KCN tested. In contrast, LED was able to completely reverse the detrimental effect of tetrodotoxin, which only indirectly down-regulated enzyme levels. Among the wavelengths tested (670, 728, 770, 830, and 880 nm), the most effective ones (670 nm and 830 nm) paralleled the NIR absorption spectrum of oxidized cytochrome c oxidase.
5.3 Nerve regeneration. Animal models have been employed to study LLLT effects in nerve repair [73, 74]. Byrnes et al. [56] used 1,600 J/cm2 of 810-nm diode laser to improve healing and functionality in a T9 dorsal hemisection of the spinal cord in rats. Anders et al. [75] studied LLLT for regenerating crushed rat facial nerves; by comparing 361, 457, 514, 633, 720, and 1064 nm, and found the best response with 162.4 J/cm2 of 633 nm HeNe laser.
6. CLINICAL STUDIES
Low-power laser therapy is used by physical therapists to treat a wide variety of acute and chronic musculoskeletal aches and pains, by dentists to treat inflamed oral tissues and to heal diverse ulcerations, by dermatologists to treat edema, non-healing ulcers, burns, and dermatitis, by orthopedists to relieve pain and treat chronic inflammations and autoimmune diseases, and by other specialists, as well as general practitioners. Laser therapy is also widely used in veterinary medicine (especially in racehorse-training centers), and in sports-medicine and rehabilitation clinics (to reduce swelling and hematoma, relieve pain, improve mobility, and treat acute soft-tissue injuries). Lasers and LEDs are applied directly to the respective areas (e.g., wounds, sites of injuries) or to various points on the body (acupuncture points, muscle-trigger points). However, one of the most important limitations to advancing the LLLT field into mainstream medical practice is the lack of appropriately controlled and blind clinical trials. The trials should be prospective, placebo controlled, and double blinded, and contain sufficient subjects to allow statistically valid conclusions to be reached.
Clinical applications of low-power laser therapy are diverse. The field is characterized by a variety of methodologies, and uses of various light sources (lasers, LEDs) with different parameters (wavelength, output power, continuous-wave or pulsed operation modes, pulse parameters). In recent years, longer wavelengths (~800 to 900 nm) and higher output powers (to 100 mW) have been preferred in therapeutic devices, especially to allow deeper tissue penetration. In 2002, MicroLight Corp received 510K FDA clearance for the ML 830 nm diode laser for the treatment of carpal tunnel syndrome. There were several controlled trials reporting significant improvement in pain, and some improvement in objective outcome measures [76-78]. Since then several light sources have been approved as equivalent to an infrared heating lamp for treating a wide-range of musculoskeletal disorders with no supporting clinical studies.
7. UNRESOLVED QUESTIONS
7.1 Wavelength. This is probably the parameter where there is most agreement in the LLLT community. Wavelengths in the 600-700 nm range are chosen for treating superficial tissue, and wavelengths between 780 and 950 nm are chosen for deeper-seated tissues, due to longer optical penetration distances through tissue. Wavelengths between 700 and 770 nm are not considered to have much activity. Some devices combine a red wavelength with a NIR wavelength on the basis that the combination of two wavelengths can have additive effects, and can also allow the device to be more broadly utilized to treat more diseases. There is of course much more work to be done to define what is the optimum wavelength for the different indications for which LLLT is employed.
7.2 Laser vs non-coherent light. One of the most topical and widely discussed issues in the LLLT clinical community is whether the coherence and monochromatic nature of laser radiation have additional benefits, as compared with more broad-band light from a conventional light source or LED with the same center wavelength and intensity. Two aspects of this problem must be distinguished: the coherence of light itself and the coherence of the interaction of light with matter (biomolecules, tissues). The latter interaction produces the phenomenon known as laser speckle, which has been postulated to play a role in the photobiomodulation interaction with cells and subcellular organelles. It is difficult to design an experiment to directly compare coherent laser light with non-coherent non-laser light for the following reason. Laser light is almost always monochromatic with a bandwidth of 1 nm or less, and it is very difficult to generate light from any other source (even an LED) that has a bandwidth narrower than 10-20 nm, therefore it will be uncertain if observed differences are due to coherent versus non-coherent light, or due to monochromatic versus narrow bandwidth light.
7.3 Dose. Because of the possible existence of a biphasic dose response curve referred to above, choosing the correct dosage of light (in terms of energy density) for any specific medical condition is difficult. In addition there has been some confusion in the literature about the delivered fluence when the light spot is small. If 5J of light is given to a spot of 5 mm2, the fluence is 100 J/cm2, which is nominally the same fluence as 100 J/cm2 delivered to 10 cm2, but the total energy delivered in the latter case is 200 times greater. The dose of light that is used depends on the pathology being treated, and in particular upon how deep the light is thought to need to penetrate into the tissue. Doses that are frequently used in the red wavelengths for fairly superficial diseases tend to be in the region of 4 J/cm2 with a range of 1-10 J/cm2. Doses of the NIR wavelengths that tend to be employed for deeper-seated disorders can be higher than these values, i.e., in the 10-50 J/cm2 range. The light treatment is usually repeated either every day or every other day, and a course of treatment can last for periods around two weeks.
7.4 Pulsed or CW. There have been some reports that pulse structure is an important factor in LLLT; for instance Ueda et al. [79, 80] found better effects using 1 or 2 Hz pulses than 8 Hz or CW 830 nm laser on rat bone cells, but the underlying mechanism for this effect is unclear.
7.5 Polarization status. There are some claims that polarized light has better effects in LLLT applications than otherwise identical non-polarized light (or even 90-degree rotated polarized light) [81]. However, it is known that polarized light is rapidly scrambled in highly scattering media such as tissue (probably in the first few hundred µm), and it therefore seems highly unlikely that polarization could play a role, except for superficial applications to the upper layers of the skin.
7.6 Systemic effects. Although LLLT is mostly applied to localized diseases and its effect is often considered to be restricted to the irradiated area, there are reports of systemic effects of LLLT acting at a site distant from the illumination [82, 83]. It is well known that UV light can have systemic effects [84], and it has been proposed that red and NIR light can also have systemic effects. These have been proposed to be mediated by soluble mediators such as endorphins and serotonin. There is a whole field known as laser acupuncture [85] in which the stimulation of specific acupuncture points by a focused laser beam is proposed to have similar effects at distant locations to the more well known needle acupuncture techniques.
EDITOR'S NOTE (02/23/13)
Follow this LINK for a list of 3080 papers on LLLT on PubMed.
REFERENCES
[1] E. Mester, B. Szende and P. Gartner, The effect of laser beams on the growth of hair in mice, Radiobiol Radiother (Berl) 9 (1968) 621-6.
[2] R. Roelandts, The history of phototherapy: something new under the sun?, J Am Acad Dermatol 46 (2002) 926-30.
[3] A.N. Pereira, P. Eduardo Cde, E. Matson and M.M. Marques, Effect of low-power laser irradiation on cell growth and procollagen synthesis of cultured fibroblasts, Lasers Surg Med 31 (2002) 263-7.
[4] J.S. Kana, G. Hutschenreiter, D. Haina and W. Waidelich, Effect of low-power density laser radiation on healing of open skin wounds in rats, Arch Surg 116 (1981) 293-6.
[5] A.P. Sommer, A.L. Pinheiro, A.R. Mester, R.P. Franke and H.T. Whelan, Biostimulatory windows in low-intensity laser activation: lasers, scanners, and NASA's light-emitting diode array system, J Clin Laser Med Surg 19 (2001) 29-33.
[6] J.C. Sutherland, Biological effects of polychromatic light, Photochem Photobiol 76 (2002) 164-70.
[7] T. Karu, Laser biostimulation: a photobiological phenomenon, J Photochem Photobiol B 3 (1989) 638-40.
[8] T.I. Karu and N.I. Afanas'eva, Cytochrome c oxidase as the primary photoacceptor upon laser exposure of cultured cells to visible and near IR-range light, Dokl Akad Nauk 342 (1995) 693-5.
[9] R.A. Capaldi, F. Malatesta and V.M. Darley-Usmar, Structure of cytochrome c oxidase, Biochim Biophys Acta 726 (1983) 135-48.
[10] I. Szundi, G.L. Liao and O. Einarsdottir, Near-infrared time-resolved optical absorption studies of the reaction of fully reduced cytochrome c oxidase with dioxygen, Biochemistry 40 (2001) 2332-9.
[11] T.I. Karu and S.F. Kolyakov, Exact action spectra for cellular responses relevant to phototherapy, Photomed Laser Surg 23 (2005) 355-61.
[12] D. Pastore, M. Greco and S. Passarella, Specific helium-neon laser sensitivity of the purified cytochrome c oxidase, Int J Radiat Biol 76 (2000) 863-70.
[13] V.G. Artyukhov, O.V. Basharina, A.A. Pantak and L.S. Sveklo, Effect of helium-neon laser on activity and optical properties of catalase, Bull Exp Biol Med 129 (2000) 537-40.
[14] W. Yu, J.O. Naim, M. McGowan, K. Ippolito and R.J. Lanzafame, Photomodulation of oxidative metabolism and electron chain enzymes in rat liver mitochondria, Photochem Photobiol 66 (1997) 866-71.
[15] S. Passarella, He-Ne laser irradiation of isolated mitochondria, J Photochem Photobiol B 3 (1989) 642-3.
[16] S.J. Ehrreich and R.F. Furchgott, Relaxation of mammalian smooth muscles by visible and ultraviolet radiation, Nature 218 (1968) 682-4.
[17] H. Chaudhry, M. Lynch, K. Schomacker, R. Birngruber, K. Gregory and I. Kochevar, Relaxation of vascular smooth muscle induced by low-power laser radiation, Photochem Photobiol 58 (1993) 661-9.
[18] R. Mittermayr, A. Osipov, C. Piskernik, S. Haindl, P. Dungel, C. Weber, Y.A. Vladimirov, H. Redl and A.V. Kozlov, Blue laser light increases perfusion of a skin flap via release of nitric oxide from hemoglobin, Mol Med 13 (2007) 22-9.
[19] L. Vladimirov, A., G.I. Klebanov, G.G. Borisenko and A.N. Osipov, Molecular and cellular mechanisms of the low intensity laser radiation effect, Biofizika 49 (2004) 339-50.
[20] Y.A. Vladimirov, A.N. Osipov and G.I. Klebanov, Photobiological principles of therapeutic applications of laser radiation, Biochemistry (Mosc) 69 (2004) 81-90.
[21] G.G. Borisenko, A.N. Osipov, K.D. Kazarinov and A. Vladimirov Yu, Photochemical reactions of nitrosyl hemoglobin during exposure to low-power laser irradiation, Biochemistry (Mosc) 62 (1997) 661-6.
[22] B. Beltran, A. Mathur, M.R. Duchen, J.D. Erusalimsky and S. Moncada, The effect of nitric oxide on cell respiration: A key to understanding its role in cell survival or death, Proc Natl Acad Sci U S A 97 (2000) 14602-7.
[23] G.C. Brown, Regulation of mitochondrial respiration by nitric oxide inhibition of cytochrome c oxidase, Biochim Biophys Acta 1504 (2001) 46-57.
[24] N. Lane, Cell biology: power games, Nature 443 (2006) 901-3.
[25] G.C. Brown and V. Borutaite, Nitric oxide inhibition of mitochondrial respiration and its role in cell death, Free Radic Biol Med 33 (2002) 1440-50.
[26] P. Ghafourifar and E. Cadenas, Mitochondrial nitric oxide synthase, Trends Pharmacol Sci 26 (2005) 190-5.
[27] E. Clementi, G.C. Brown, N. Foxwell and S. Moncada, On the mechanism by which vascular endothelial cells regulate their oxygen consumption, Proc Natl Acad Sci U S A 96 (1999) 1559-62.
[28] F. Antunes, A. Boveris and E. Cadenas, On the mechanism and biology of cytochrome oxidase inhibition by nitric oxide, Proc Natl Acad Sci U S A 101 (2004) 16774-9.
[29] T.I. Karu, L.V. Pyatibrat and N.I. Afanasyeva, Cellular effects of low power laser therapy can be mediated by nitric oxide, Lasers Surg Med 36 (2005) 307-14.
[30] V. Borutaite, A. Budriunaite and G.C. Brown, Reversal of nitric oxide-, peroxynitrite- and S-nitrosothiol-induced inhibition of mitochondrial respiration or complex I activity by light and thiols, Biochim Biophys Acta 1459 (2000) 405-12.
[31] G.A. Guzzardella, M. Fini, P. Torricelli, G. Giavaresi and R. Giardino, Laser stimulation on bone defect healing: an in vitro study, Lasers Med Sci 17 (2002) 216-20.
[32] H. Tuby, L. Maltz and U. Oron, Modulations of VEGF and iNOS in the rat heart by low level laser therapy are associated with cardioprotection and enhanced angiogenesis, Lasers Surg Med 38 (2006) 682-8.
[33] Y. Moriyama, E.H. Moriyama, K. Blackmore, M.K. Akens and L. Lilge, In vivo study of the inflammatory modulating effects of low-level laser therapy on iNOS expression using bioluminescence imaging, Photochem Photobiol 81 (2005) 1351-5.
[34] M.C. Leung, S.C. Lo, F.K. Siu and K.F. So, Treatment of experimentally induced transient cerebral ischemia with low energy laser inhibits nitric oxide synthase activity and up-regulates the expression of transforming growth factor-beta 1, Lasers Surg Med 31 (2002) 283-8.
[35] H. Friedmann, R. Lubart, I. Laulicht and S. Rochkind, A possible explanation of laser-induced stimulation and damage of cell cultures, J Photochem Photobiol B 11 (1991) 87-91.
[36] M. Eichler, R. Lavi, A. Shainberg and R. Lubart, Flavins are source of visible-light-induced free radical formation in cells, Lasers Surg Med 37 (2005) 314-9.
[37] K. Plaetzer, T. Kiesslich, B. Krammer and P. Hammerl, Characterization of the cell death modes and the associated changes in cellular energy supply in response to AlPcS4-PDT, Photochem Photobiol Sci 1 (2002) 172-7.
[38] R. Lubart, M. Eichler, R. Lavi, H. Friedman and A. Shainberg, Low-energy laser irradiation promotes cellular redox activity, Photomed Laser Surg 23 (2005) 3-9.
[39] R. Duan, T.C. Liu, Y. Li, H. Guo and L.B. Yao, Signal transduction pathways involved in low intensity He-Ne laser-induced respiratory burst in bovine neutrophils: a potential mechanism of low intensity laser biostimulation, Lasers Surg Med 29 (2001) 174-8.
[40] F.Q. Schafer and G.R. Buettner, Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple, Free Radic Biol Med 30 (2001) 1191-212.
[41] H. Liu, R. Colavitti, Rovira, II and T. Finkel, Redox-dependent transcriptional regulation, Circ Res 97 (2005) 967-74.
[42] M. Yang, N.B. Nazhat, X. Jiang, S.M. Kelsey, D.R. Blake, A.C. Newland and C.J. Morris, Adriamycin stimulates proliferation of human lymphoblastic leukaemic cells via a mechanism of hydrogen peroxide (H2O2) production, Br J Haematol 95 (1996) 339-44.
[43] W.G. Kirlin, J. Cai, S.A. Thompson, D. Diaz, T.J. Kavanagh and D.P. Jones, Glutathione redox potential in response to differentiation and enzyme inducers, Free Radic Biol Med 27 (1999) 1208-18.
[44] S. Alaluf, H. Muir-Howie, H.L. Hu, A. Evans and M.R. Green, Atmospheric oxygen accelerates the induction of a post-mitotic phenotype in human dermal fibroblasts: the key protective role of glutathione, Differentiation 66 (2000) 147-55.
[45] T. Karu, Primary and secondary mechanisms of action of visible to near-IR radiation on cells, J Photochem Photobiol B 49 (1999) 1-17.
[46] O. Tiphlova and T. Karu, Action of low-intensity laser radiation on Escherichia coli, Crit Rev Biomed Eng 18 (1991) 387-412.
[47] T.I. Karu, L.V. Pyatibrat, G.S. Kalendo and R.O. Esenaliev, Effects of monochromatic low-intensity light and laser irradiation on adhesion of HeLa cells in vitro, Lasers Surg Med 18 (1996) 171-7.
[48] P. Moore, T.D. Ridgway, R.G. Higbee, E.W. Howard and M.D. Lucroy, Effect of wavelength on low-intensity laser irradiation-stimulated cell proliferation in vitro, Lasers Surg Med 36 (2005) 8-12.
[49] D. Hawkins and H. Abrahamse, Biological effects of helium-neon laser irradiation on normal and wounded human skin fibroblasts, Photomed Laser Surg 23 (2005) 251-9.
[50] H.S. Yu, C.S. Wu, C.L. Yu, Y.H. Kao and M.H. Chiou, Helium-neon laser irradiation stimulates migration and proliferation in melanocytes and induces repigmentation in segmental-type vitiligo, J Invest Dermatol 120 (2003) 56-64.
[51] S. Young, P. Bolton, M. Dyson, W. Harvey and C. Diamantopoulos, Macrophage responsiveness to light therapy, Lasers Surg Med 9 (1989) 497-505.
[52] Y. Fujimaki, T. Shimoyama, Q. Liu, T. Umeda, S. Nakaji and K. Sugawara, Low-level laser irradiation attenuates production of reactive oxygen species by human neutrophils, J Clin Laser Med Surg 21 (2003) 165-70.
[53] Y.S. Chen, S.F. Hsu, C.W. Chiu, J.G. Lin, C.T. Chen and C.H. Yao, Effect of low-power pulsed laser on peripheral nerve regeneration in rats, Microsurgery 25 (2005) 83-9.
[54] M. Miloro, L.E. Halkias, S. Mallery, S. Travers and R.G. Rashid, Low-level laser effect on neural regeneration in Gore-Tex tubes, Oral Surg Oral Med Oral Pathol Oral Radiol Endod 93 (2002) 27-34.
[55] P. Balaban, R. Esenaliev, T. Karu, E. Kutomkina, V. Letokhov, A. Oraevsky and N. Ovcharenko, He-Ne laser irradiation of single identified neurons, Lasers Surg Med 12 (1992) 329-37.
[56] K.R. Byrnes, R.W. Waynant, I.K. Ilev, X. Wu, L. Barna, K. Smith, R. Heckert, H. Gerst and J.J. Anders, Light promotes regeneration and functional recovery and alters the immune response after spinal cord injury, Lasers Surg Med 36 (2005) 171-85.
[57] S.O. el Sayed and M. Dyson, Effect of laser pulse repetition rate and pulse duration on mast cell number and degranulation, Lasers Surg Med 19 (1996) 433-7.
[58] R.A. Lopes-Martins, R. Albertini, P.S. Martins, J.M. Bjordal and H.C. Faria Neto, Spontaneous effects of low-level laser therapy (650 nm) in acute inflammatory mouse pleurisy induced by Carrageenan, Photomed Laser Surg 23 (2005) 377-81.
[59] A.D. Agaiby, L.R. Ghali, R. Wilson and M. Dyson, Laser modulation of angiogenic factor production by T-lymphocytes, Lasers Surg Med 26 (2000) 357-63.
[60] S. Passarella, E. Casamassima, S. Molinari, D. Pastore, E. Quagliariello, I.M. Catalano and A. Cingolani, Increase of proton electrochemical potential and ATP synthesis in rat liver mitochondria irradiated in vitro by helium-neon laser, FEBS Lett 175 (1984) 95-9.
[61] M. Greco, G. Guida, E. Perlino, E. Marra and E. Quagliariello, Increase in RNA and protein synthesis by mitochondria irradiated with helium-neon laser, Biochem Biophys Res Commun 163 (1989) 1428-34.
[62] D. Pastore, M. Greco, V.A. Petragallo and S. Passarella, Increase in H+/e- ratio of the cytochrome c oxidase reaction in mitochondria irradiated with helium-neon laser, Biochem Mol Biol Int 34 (1994) 817-26.
[63] Y. Zhang, S. Song, C.C. Fong, C.H. Tsang, Z. Yang and M. Yang, cDNA microarray analysis of gene expression profiles in human fibroblast cells irradiated with red light, J Invest Dermatol 120 (2003) 849-57.
[64] R.F. Lyons, R.P. Abergel, R.A. White, R.M. Dwyer, J.C. Castel and J. Uitto, Biostimulation of wound healing in vivo by a helium-neon laser, Ann Plast Surg 18 (1987) 47-50.
[65] H.S. Yu, K.L. Chang, C.L. Yu, J.W. Chen and G.S. Chen, Low-energy helium-neon laser irradiation stimulates interleukin-1 alpha and interleukin-8 release from cultured human keratinocytes, J Invest Dermatol 107 (1996) 593-6.
[66] V.K. Poon, L. Huang and A. Burd, Biostimulation of dermal fibroblast by sublethal Q-switched Nd:YAG 532 nm laser: collagen remodeling and pigmentation, J Photochem Photobiol B 81 (2005) 1-8.
[67] N. Kipshidze, V. Nikolaychik, M.H. Keelan, L.R. Shankar, A. Khanna, R. Kornowski, M. Leon and J. Moses, Low-power helium: neon laser irradiation enhances production of vascular endothelial growth factor and promotes growth of endothelial cells in vitro, Lasers Surg Med 28 (2001) 355-64.
[68] A. Khanna, L.R. Shankar, M.H. Keelan, R. Kornowski, M. Leon, J. Moses and N. Kipshidze, Augmentation of the expression of proangiogenic genes in cardiomyocytes with low dose laser irradiation in vitro, Cardiovasc Radiat Med 1 (1999) 265-9.
[69] A.R. Medrado, L.S. Pugliese, S.R. Reis and Z.A. Andrade, Influence of low level laser therapy on wound healing and its biological action upon myofibroblasts, Lasers Surg Med 32 (2003) 239-44.
[70] E.J. Neiburger, Rapid healing of gingival incisions by the helium-neon diode laser, J Mass Dent Soc 48 (1999) 8-13, 40.
[71] J.T. Eells, M.M. Henry, P. Summerfelt, M.T. Wong-Riley, E.V. Buchmann, M. Kane, N.T. Whelan and H.T. Whelan, Therapeutic photobiomodulation for methanol-induced retinal toxicity, Proc Natl Acad Sci U S A 100 (2003) 3439-44.
[72] M.T. Wong-Riley, H.L. Liang, J.T. Eells, B. Chance, M.M. Henry, E. Buchmann, M. Kane and H.T. Whelan, Photobiomodulation directly benefits primary neurons functionally inactivated by toxins: role of cytochrome c oxidase, J Biol Chem 280 (2005) 4761-71.
[73] D. Gigo-Benato, S. Geuna and S. Rochkind, Phototherapy for enhancing peripheral nerve repair: a review of the literature, Muscle Nerve 31 (2005) 694-701.
[74] J.J. Anders, S. Geuna and S. Rochkind, Phototherapy promotes regeneration and functional recovery of injured peripheral nerve, Neurol Res 26 (2004) 233-9.
[75] J.J. Anders, R.C. Borke, S.K. Woolery and W.P. Van de Merwe, Low power laser irradiation alters the rate of regeneration of the rat facial nerve, Lasers Surg Med 13 (1993) 72-82.
[76] K. Branco and M.A. Naeser, Carpal tunnel syndrome: clinical outcome after low-level laser acupuncture, microamps transcutaneous electrical nerve stimulation, and other alternative therapies--an open protocol study, J Altern Complement Med 5 (1999) 5-26.
[77] J. Irvine, S.L. Chong, N. Amirjani and K.M. Chan, Double-blind randomized controlled trial of low-level laser therapy in carpal tunnel syndrome, Muscle Nerve 30 (2004) 182-7.
[78] M.I. Weintraub, Noninvasive laser neurolysis in carpal tunnel syndrome, Muscle Nerve 20 (1997) 1029-31.
[79] Y. Ueda and N. Shimizu, Pulse irradiation of low-power laser stimulates bone nodule formation, J Oral Sci 43 (2001) 55-60.
[80] Y. Ueda and N. Shimizu, Effects of pulse frequency of low-level laser therapy (LLLT) on bone nodule formation in rat calvarial cells, J Clin Laser Med Surg 21 (2003) 271-7.
[81] M.S. Ribeiro, F. Da Silva Dde, C.E. De Araujo, S.F. De Oliveira, C.M. Pelegrini, T.M. Zorn and D.M. Zezell, Effects of low-intensity polarized visible laser radiation on skin burns: a light microscopy study, J Clin Laser Med Surg 22 (2004) 59-66.
[82] T. Moshkovska and J. Mayberry, It is time to test low level laser therapy in Great Britain, Postgrad Med J 81 (2005) 436-41.
[83] L.A. Santana-Blank, E. Rodriguez-Santana and K.E. Santana-Rodriguez, Photo-infrared pulsed bio-modulation (PIPBM): a novel mechanism for the enhancement of physiologically reparative responses, Photomed Laser Surg 23 (2005) 416-24.
[84] M.L. Kripke, Ultraviolet radiation and immunology: something new under the sun--presidential address, Cancer Res 54 (1994) 6102-5.
[85] P. Whittaker, Laser acupuncture: past, present, and future, Lasers Med Sci 19 (2004) 69-80.
8/14/08